
研究の背景
アポトーシスとは
アポトーシスとは、1972年にイギリスのKerrとWyllieによって命名された細胞死の形態の一つである。細胞が膨潤して崩壊していくネクローシス(壊死)とは異なり、アポトーシスを起こした細胞は、その内容物を含んだまま隣接する食細胞にとりこまれて処理されるため、『クリーンな細胞死』ともいわれる。
アポトーシスの分子機構
アポトーシスは"death factor"として作用するサイトカイン(Fasリガンド) とその受容体(Fas)を介したシグナル伝達系の活性化によって惹起される(Cell 88, 355, 1997)。
Fas,Fasリガンド遺伝子に変異を持つマウス(lprマウス、gldマウス)では、リンパ球の異常な蓄積や、重篤な自己免疫疾患の誘発が見られる事から、この細胞死の機構が生体の恒常性維持に必須の役割を果たす事が明らかにされてきた [これまでの研究 "Fas-induced apoptosis"の項]。
DNAの断片化の分子機構
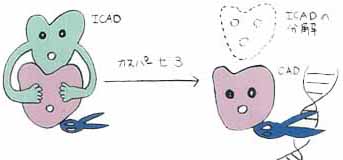
我々の研究室では、Fasリガンドによるアポトーシスのシグナル伝達機構の解析から、アポトーシス細胞に見られるDNAの断片化が、カスパーゼによって活性化されるCAD(caspase-activated DNase)というDNA分解酵素によって引き起こされる事を示した (Nature 391, 43, 1998: Nature 391, 96, 1998)。[これまでの研究 "DNA fragmentation by CAD during apoptosis"の項]
さらに、CADノックアウトマウスの樹立・解析から、このマウスから調製した細胞を試験管内で培養しアポトーシスを誘導しても染色体DNAの分解はおこらないが、動物の生体内でアポトーシスを起こしている細胞を観察すると、CAD遺伝子が欠損している細胞でもDNAの分解は正常に起こっている事がわかった。しかもこのDNAの分解はマクロファージに貪食された細胞でのみ見られた。この事から、アポトーシスを起こした細胞はマクロファージによって速やかにとりこまれ、マクロファージがアポトーシス細胞のDNAを分解する可能性が示唆された。[これまでの研究 "Macrophage-mediated DNA fragmentation"の項]
このことを確かめるために、マクロファージに存在するDNA分解酵素(DNaseII)遺伝子のノックアウトマウスを作成した。その結果、DNaseIIノックマウスは胎生致死に陥り全く生まれてこないという意外な事実が明らかとなった。[最近の研究 "DNase IIノックアウトマウス"の項]
ところで、染色体DNAの分解を担っている酵素にはCAD,DNaseIIの他にDLAD(DNaseII-like acid DNase)がある。DLADの組織における発現を調べた所、肝臓に加えて目のレンズにおいて非常に強い発現が見られた。目のレンズ細胞には核、ミトコンドリア、小胞体などのオルガネラが存在せず、オルガネラの消失が水晶体の透明性を維持するために必要と考えられていたが、その分子機構は全く不明であった。そこで当研究室では水晶体・繊維細胞でのDNA分解の機構を明らかにするため、DLADノックアウトマウスを作成した。[最近の研究 "DLADノックアウトマウス"の項]
ファゴサイトーシスとは?
アポトーシス細胞はマクロファージや好中球などの食細胞に速やかに貪食処理(ファゴサイトーシス)される。この貪食処理の仕組みによって、死細胞から炎症を引き起こす分子の放出を防いでいる。マクロファージはアポトーシス細胞を貪食するが、生きている細胞を貪食することはない。このメカニズムは、アポトーシス細胞が細胞表面に、"eat me" シグナルをマクロファージに対して提示し、これをマクロファージが認識し、貪食しているためだと考えられている。
当研究室では、マクロファージによるアポトーシス細胞特異的な認識機構に重要な役割を果たす分子として、MFG-E8(Milk Fat Globule EGF-factor 8)というタンパク質を同定した。MFG-E8は活性化されたマクロファージによって分泌されると、マクロファージ上に発現するインテグリンと、アポトーシス細胞上に提示されるeat meシグナル(ホスファチジルセリン)の両方に結合し、二つの細胞を橋渡しをする事で、特異的な貪食を促進する事が示された。[最近の研究 "マクロファージによるアポトーシス細胞の貪食機構"の項]
![]()